The quest to understand our origins has been a topic that has kept many paleoanthropologists engaged in reconstructing the evolutionary tree in the past few years. Fossil specimens from different sites in the world have been the primary source of data, most of which are from the Late Miocene period (11.6-5.3mya), and the Plio-pleistocene period (5.3mya-11kya). In tracing our evolutionary lineage, we understand that humans belong to the superfamily Hominoidea, which has two extant families: hylobatidae/lesser apes made up of the siamangs and gibbons inhabiting the forested areas in Southeast Asia, and the hominidae/greater apes, that include orangutans (found in Borneo and Sumatra), the African apes (gorillas, bonobos, chimpanzees), and humans.
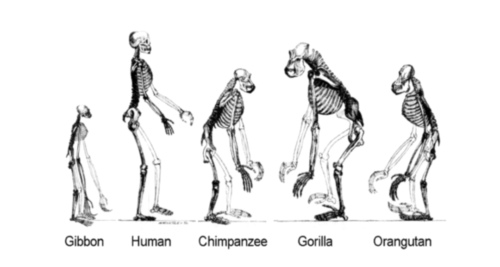
Members of the superfamily Hominoidea. The differences in the forelimbs and hindlimbs shows the different locomotory adaptations. The orangutan and gibbon are adapted to a fully arboreal lifestyle, the chimp and gorilla are quadrupedal knuckle-walkers with occasional bipedalism, while the human is habitually bipedal.
Based on studies of DNA, bonobos and chimpanzees are moreclosely related to humans than either is to gorillas. It is likely that sometime back in history we shared an ancestry. Genetic data shows that we shared a last common ancestor with the chimpanzee about 7 mya. Overtime, we have used chimps as a reference in evolutionary reconstruction, mainly because of the morphological and behavioural resemblance we share, which has led some scientists proposing a chimp-like common ancestor. However, it still remains a debatable topic, provided there are currently no fossil chimps, we can not say that chimps have remained unchanged in the past 7 million years.
Although various theories exist, they fall short in explaining the origins of bipedalism, and therefore skeletal remains from hominins have been carefully studied to understand when, how, and why bipedalism might have started as a preferred mode of locomotion. The earliest hominins (Sahelanthropus tchadensis, Orrorin tugenensis, Ardipithecus ramidus) presents us with a mosaic of derived and primitive features in the cranium, dentition, and the few postcranial elements. The trend seen in reduction of the canines from the earliest hominins, through the australopithecines, and most pronounced in Homo, gives us more insight about our evolutionary trajectory. The dental-plan, especially in canines, has been used to study the paleo-diets and social organization of our ancestors. Other morphological changes are seen in the cranium alongside facial prognathism. Increased brain size and changing diets, especially consuming softer foods, would have contributed to the reduction of the teeth and jaws, and thus, reduced facial prognathism. An interesting aspect of jaw and teeth reduction could also have played a significant role in chin evolution in Homo sapiens. To understand the challenges most paleoanthropologists face while describing and sorting out specimens, students have been engaging in discussions on the various morphological changes, tracing the developmental changes, and also exercising on creating phylogenetic trees of some hypothetical species as assigned by Dr. Mongle.
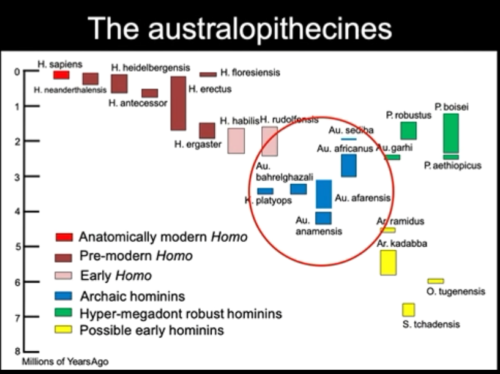
The temporal range of australopithecines.
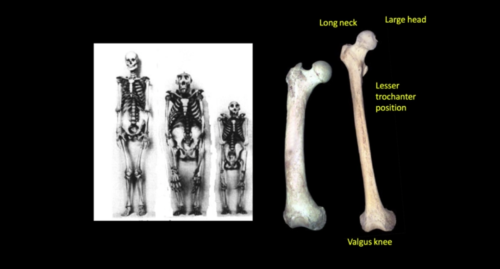
Bipedalism in humans differs from apes mainly by having a long femur with a long neck as compared to the short femur and neck in chimps. In addition, the alignment of the femur brings about a valgus angle at the knee in humans, where the distal ends of the femur are much closer as compared to the proximal ends.
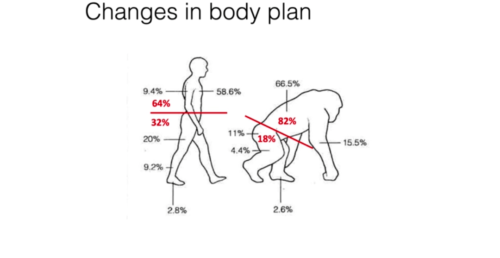
An evolutionary advantage of bipedalism is energy efficiency, thus favouring long distance movement.
While research is ongoing to understand our origins, it is correct to say that our evolutionary tree has become more bushy. Identifiable morphological, behavioral, and spatio-temporal scales have been used to make inferences parsimoniously on our complex evolutionary tree. However, we still see contemporaneity of species in the record, paving way for more research on major changes in species diversity and landscape exploitation.




